1 A água no sistema solo-planta-atmosfera
O sistema solo-planta-atmosfera (SPA) é um sistema complexo e interativo que regula o movimento da água, nutrientes e energia do solo para a atmosfera, passando pela planta. A água, elemento essencial para a vida das plantas, é absorvida pelas raízes, transportada através da planta e, finalmente, evaporada pelas folhas no processo conhecido como transpiração.
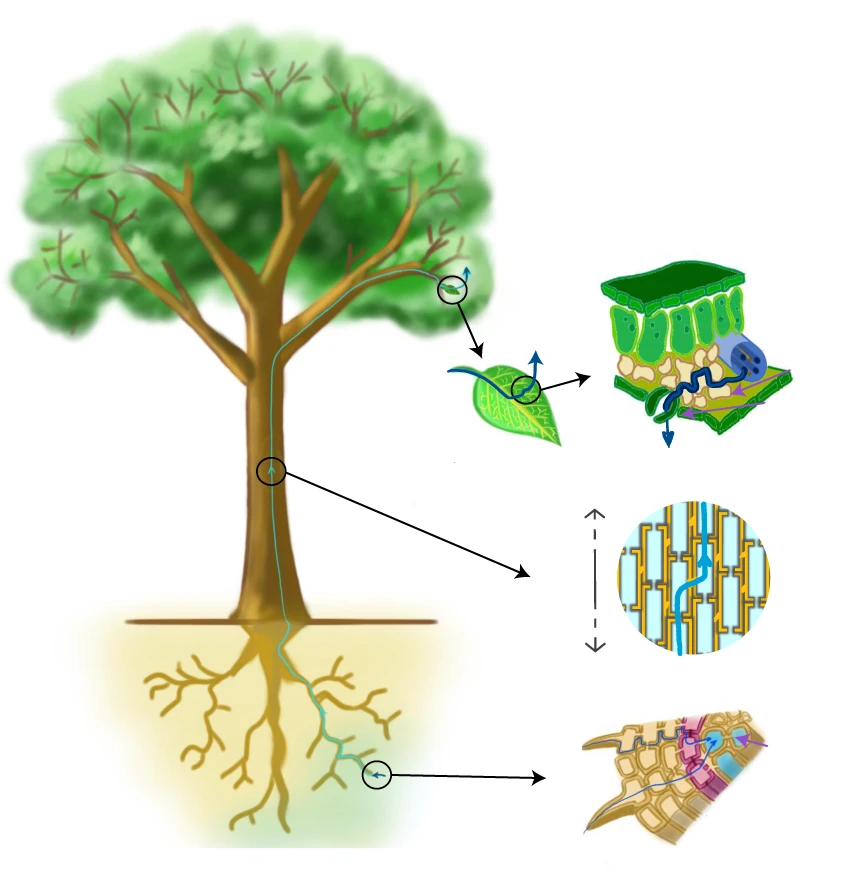
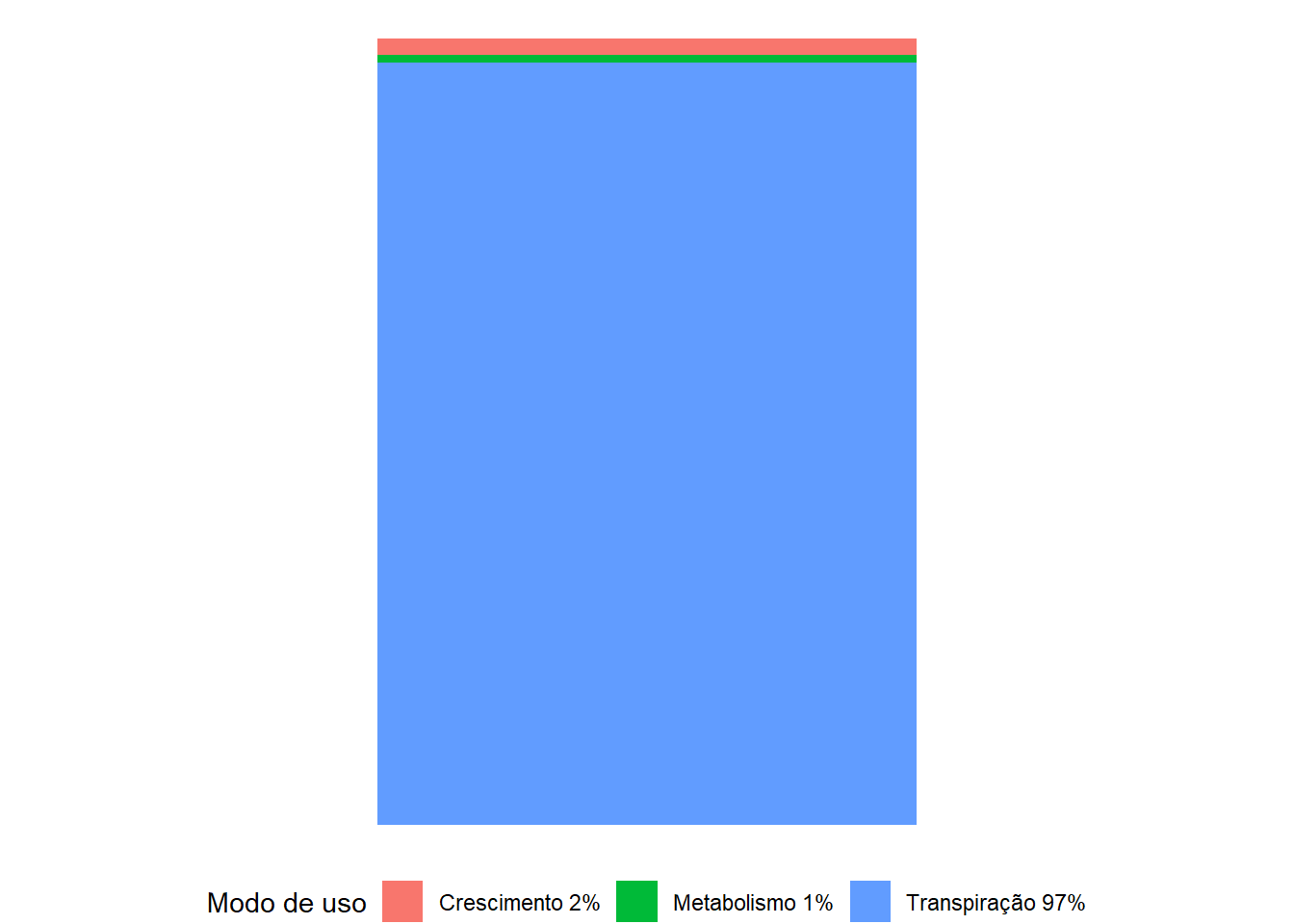
A maior parte da água absorvida pelas plantas é usada na transpiração (cerca de 97%), que compreende a absorção pelas raízes, transporte pela planta e evaporação nas superfícies foliares (Figura 1.2). Apenas uma pequena quantidade permanece na planta, no crescimento vegetal (cerca de 2%) e nas reações bioquímicas da fotossíntese e em outros processos metabólicos (cerca de 1%).
Árvores pequenas (2-3 metros de altura), podem precisar de cerca de 20 a 40 litros de água por dia. Árvores de médio porte (4-7 metros de altura), podem necessitar de aproximadamente 40 a 100 litros de água por dia. Já árvores grandes (acima de 10 metros de altura), como carvalhos ou eucaliptos, podem precisar de 100 a 200 litros ou mais por dia, especialmente durante os meses quentes de verão.
1.1 O Papel Fisiológico da Transpiração
A transpiração, classicamente definida como a perda de água pelas superfícies vegetais na forma de vapor, é um processo essencial para as plantas, com múltiplas funções que contribuem para o seu crescimento, desenvolvimento e adaptação ao ambiente. O entendimento da importância da transpiração é fundamental para o manejo adequado da água na agricultura, garantindo a produtividade das culturas e o uso eficiente dos recursos hídricos. Nesse sentido, a manutenção desse fluxo de água ao longo do sistema solo-planta-atmosfera é responsável por impulsionar processos vitais, incluindo:
Manutenção da turgescência e integridade estrutural: A água contida nos vacúolos celulares exerce a pressão de turgor, força mecânica fundamental para conferir rigidez física, especialmente em tecidos não lenhosos. É essa pressão contínua que sustenta a arquitetura espacial da planta para a captação de luz; sua perda direta resulta no murchamento imediato de folhas e hastes.
Transporte de água e nutrientes: A transpiração é o “motor” do sistema condutor. O fluxo transpiratório cria um gradiente de tensão que impulsiona a água das raízes até o dossel. Esse fluxo de massa é o veículo que garante a absorção e a distribuição eficiente de nutrientes minerais do solo por toda a planta, suprindo as demandas essenciais para o crescimento.
Termorregulação foliar: A mudança de fase da água, de líquida para vapor, exige o consumo de energia na forma de calor latente de vaporização. Esse processo atua como um sistema de resfriamento ativo, dissipando a energia térmica acumulada nas folhas. Esse resfriamento pode abaixar a temperatura da folha em 10°C a 15°C em relação à temperatura ambiente , prevenindo a desnaturação de enzimas termossensíveis vitais. Um exemplo clássico é a RuBisCo, cuja temperatura ideal de funcionamento fica entre 25°C e 30°C, mas que poderia atingir até 60°C sob sol pleno se não fosse o efeito termorregulador da transpiração.
No entanto, a função primordial e mais importante da transpiração é viabilizar as trocas gasosas. Para que a planta realize a fotossíntese e acumule biomassa, é imprescindível a absorção de dióxido de carbono (C2) da atmosfera. Essa captação ocorre por meio da abertura dos estômatos, que operam como válvulas de comunicação entre o interior da folha e o ambiente externo.
O grande desafio imposto pelo ambiente terrestre é que a atmosfera atua como um dreno constante, sendo quase sempre mais seca que o interior das folhas. Dessa forma, a perda de vapor de água para a atmosfera é um efeito colateral físico incontornável da abertura estomática. Ou seja, enquanto o CO₂ entra, a água inevitavelmente sai. A planta vive, portanto, um constante balanço entre a aquisição de carbono e a perda de água.O grande desafio imposto pelo ambiente terrestre é que a atmosfera atua como um dreno constante, sendo quase sempre mais seca que o interior das folhas. Dessa forma, a perda de vapor de água para a atmosfera é um efeito colateral físico incontornável da abertura estomática. Ou seja, enquanto o CO2 entra, a água inevitavelmente sai. A planta vive, portanto, um constante balanço entre a aquisição de carbono e a perda de água.
A magnitude desse “custo hídrico” varia consideravelmente entre as espécies vegetais, dependendo estritamente do seu mecanismo fotossintético. Essa relação é quantificada pela razão transpiratória, definida como a massa de água perdida pela planta para cada unidade de massa de matéria seca produzida. As diferenças de eficiência no uso da água são notáveis e dividem as plantas em três grandes grupos:
Plantas C3: Culturas como soja, feijão e trigo apresentam o maior gasto hídrico, transpirando de 450 a 950 gramas de água para cada grama de matéria seca acumulada.
Plantas C4: Culturas como milho e sorgo são fisiologicamente mais eficientes no uso da água, perdendo entre 250 e 350 gramas de água por grama de matéria seca.
Plantas MAC (Metabolismo Ácido das Crassuláceas): Espécies adaptadas à aridez extrema (como palma forrageira e abacaxi) abrem seus estômatos à noite, perdendo apenas de 50 a 160 gramas de água por grama de matéria seca produzida.
1.2 Movimento da água no SPA
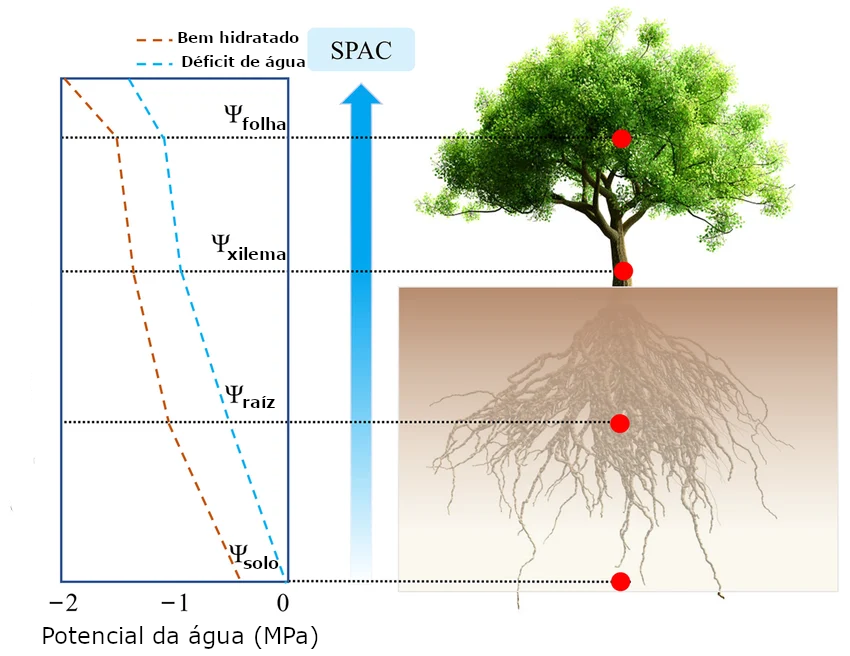
O movimento da água no sistema solo-planta-atmosfera (SPA) é um processo passivo, impulsionado pela diferença de potencial hídrico entre o solo, a planta e a atmosfera. A água flui de regiões com maior potencial hídrico (maior energia potencial) para regiões com menor potencial hídrico (menor energia potencial).
A energia da radiação solar é a força motriz do movimento da água no SPA. A radiação solar aquece a superfície do solo e das plantas, aumentando a taxa de evapotranspiração. A evapotranspiração, por sua vez, diminui o potencial hídrico nas folhas, criando um gradiente de potencial que impulsiona o fluxo de água do solo para as raízes, das raízes para as folhas e das folhas para a atmosfera.
O fluxo de calor latente, que representa a energia utilizada na evaporação da água, corresponde de 80% a 90% do saldo de radiação, o que demonstra a importância da transpiração no balanço energético do sistema SPA.
A Figura 1.4 ilustra o gradiente de potencial hídrico no sistema SPA. O potencial hídrico é mais alto no solo e diminui progressivamente à medida que a água se move em direção à atmosfera. A maior diferença de potencial hídrico ocorre entre a folha e a atmosfera, o que impulsiona a transpiração.
1.2.1 Movimento da água do solo para a raiz
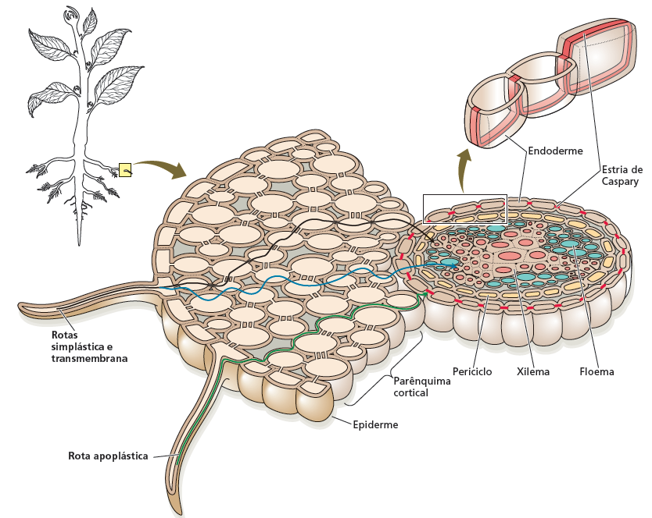
O movimento da água do solo para a raiz é um processo complexo que envolve diferentes rotas de transporte e mecanismos de regulação. A estrutura da raiz, com seus pelos radiculares e a endoderme com estrias de Caspary, permite que a planta absorva água e nutrientes do solo de forma eficiente e controlada, garantindo o suprimento adequado para o crescimento e desenvolvimento.
A zona de absorção, localizada nas extremidades das raízes, é a principal região responsável pela captação de água e nutrientes do solo. Os pelos radiculares, que são extensões das células epidérmicas, aumentam significativamente a área de superfície das raízes, maximizando a absorção.
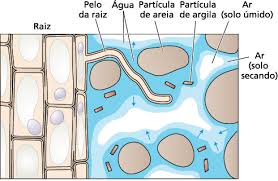
A água absorvida pelos pelos radiculares precisa atravessar as camadas celulares da raiz para chegar ao xilema, o tecido vascular responsável pelo transporte de água e nutrientes para as demais partes da planta.
A Figura 1.6 fornece uma representação detalhada da anatomia da raiz e das vias de transporte de água. Existem três rotas principais pelas quais a água pode se mover do solo para o xilema:
Rota apoplástica: A água se move através das paredes celulares e dos espaços intercelulares, sem atravessar as membranas celulares. Essa rota é mais rápida, mas menos seletiva, permitindo a passagem de água e solutos dissolvidos.
Rota simplástica: A água se move através dos plasmodesmos, que são canais que conectam os citoplasmas das células adjacentes. Essa rota é mais lenta, mas mais seletiva, permitindo o controle da passagem de água e solutos.
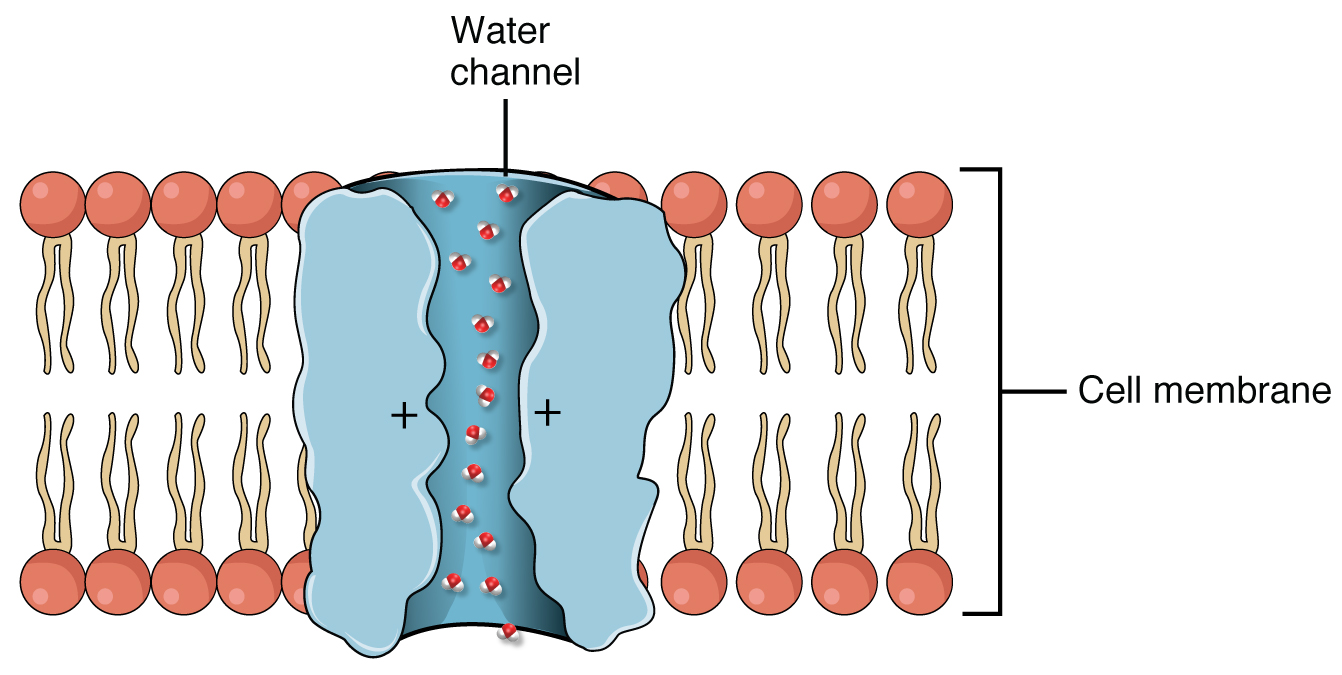
Rota transmembrana: A água atravessa as membranas celulares, entrando e saindo das células. Essa rota é auxiliada pelas aquaporinas, proteínas que facilitam o transporte de água através das membranas. A rota transmembrana é importante para o transporte de água, mas menos utilizada para o transporte de solutos.
As aquaporinas (Figura 1.7) são proteínas integrais de membrana que formam canais seletivos estruturados especificamente para a passagem de água. Embora as moléculas de água consigam atravessar a bicamada lipídica das células vegetais de forma independente por difusão simples, esse processo é consideravelmente lento. As aquaporinas contornam essa limitação ao permitir a difusão linear das moléculas de água através de seus poros, acelerando enormemente o transporte hídrico entre os meios extracelular e intracelular.
A endoderme (Figura 1.6), sendo a última camada celular que envolve o cilindro vascular antes do xilema, desempenha um papel na regulação do movimento da água e dos nutrientes do solo para a planta. As células da endoderme possuem as estrias de Caspary, que são faixas nas paredes celulares impregnadas com suberina, um material altamente hidrofóbico e impermeável.
Essas estrias impedem o fluxo livre de água e solutos pela rota apoplástica. Como essa via extracelular é fisicamente bloqueada, toda a água e os íons dissolvidos, sem exceção, são forçados a cruzar a membrana plasmática, entrando na rota simplástica ou transmembrana, para conseguir chegar ao xilema.
Essa transição obrigatória é uma etapa de controle essencial. Ao forçar a passagem pelo interior da célula (simplasto), a membrana plasmática atua como um filtro altamente seletivo. Isso garante a seletividade do processo, permitindo que a planta controle ativamente quais substâncias ela absorve do solo e quais ela impede de entrar no sistema vascular. Trata-se de um mecanismo biológico de segurança vital, evitando elementos prejudiciais,como metais pesados (chumbo, cádmio), excesso de sais e poluentes ambientais (como resíduos de pesticidas ou herbicidas). Além disso, a endoderme regula rigorosamente o fluxo de micronutrientes essenciais, como ferro (Fe), manganês (Mn) e zinco (Zn), que são imprescindíveis em pequenas quantidades, mas que se tornariam tóxicos caso se acumulassem em altas concentrações na planta.
1.2.2 Movimento da água no xilema
Uma vez que a água transpõe as barreiras da raiz e atinge o cilindro vascular, seu fluxo passa a ocorrer quase exclusivamente pelo xilema, que constitui mais de 99% de todo o trajeto hídrico na planta. Esse movimento ascendente é um processo físico complexo, dependente tanto da estrutura anatômica especializada desse tecido condutor quanto da interação de diferentes forças de tração. Funcionando como uma rede contínua de tubos microscópicos, o xilema é o grande responsável por transportar a solução do solo contra a força da gravidade, conectando o sistema radicular às folhas, onde a transpiração atuará como a principal força motriz. Essa ascensão ininterrupta é fundamental para o crescimento e o desenvolvimento vegetal, garantindo a distribuição eficiente do suprimento hídrico e de nutrientes minerais para todas as células da parte aérea.
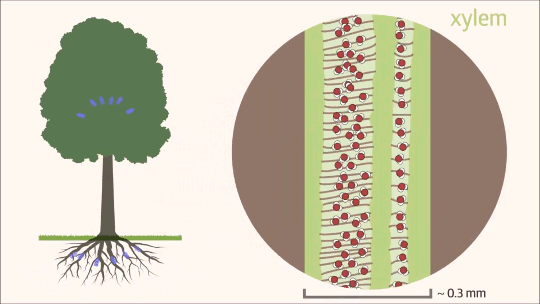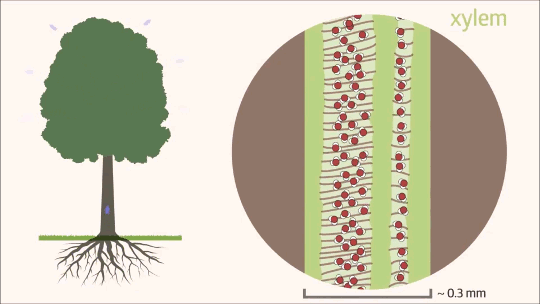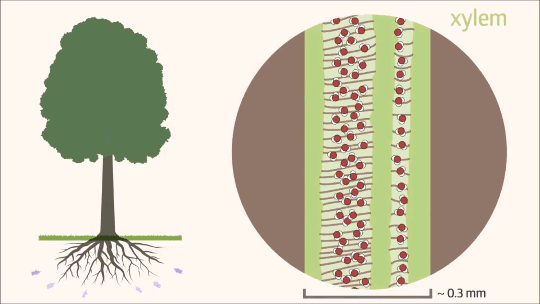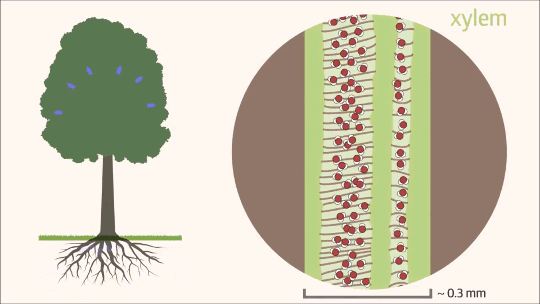
O mecanismo físico que explica a ascensão contínua da água contra a gravidade, desde as raízes até as folhas mais altas de uma árvore, é descrito pela Teoria da Coesão-Tensão. É importante destacar que esse processo não exige gasto de energia metabólica por parte da planta para bombear o fluido. Trata-se de um movimento estritamente passivo, impulsionado pela atmosfera e sustentado pela interação de três forças físicas essenciais.
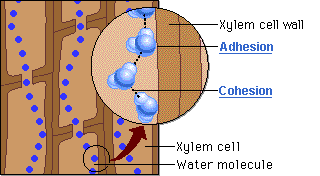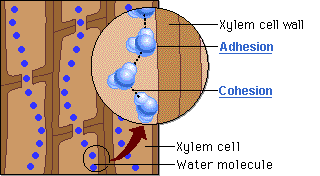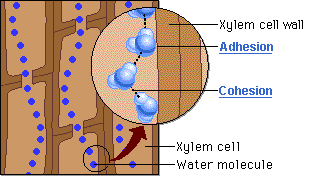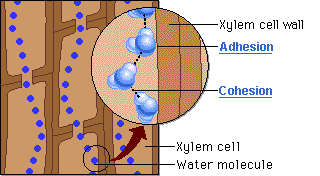
O motor primário desse movimento não está na base, mas sim no topo do sistema. A perda de água na forma de vapor pela transpiração foliar gera uma pressão negativa, ou seja, uma sucção na parte aérea. É essa tensão severa que puxa continuamente a coluna de água para cima a fim de repor o volume evaporado. Contudo, para que a tensão puxe a água sem que o fluxo se rompa, o líquido precisa agir como uma corda ininterrupta. Isso é possível graças à coesão, promovida pelas pontes de hidrogênio que mantêm as moléculas de água firmemente unidas umas às outras. Essa forte ligação intermolecular cria uma coluna líquida contínua e incrivelmente resistente à tração, capaz de suportar o estiramento sem se romper.
Ainda assim, como o peso dessa coluna de água é considerável, a coesão por si só não seria suficiente para evitar o seu colapso sob o efeito da gravidade. Nesse ponto atua a adesão, que é a notável afinidade das moléculas de água pelas paredes hidrofílicas internas do xilema, ricas em celulose e lignina. Essa aderência funciona como um suporte físico indispensável ao longo de todo o trajeto, ajudando a sustentar o peso da coluna líquida contra a força gravitacional.
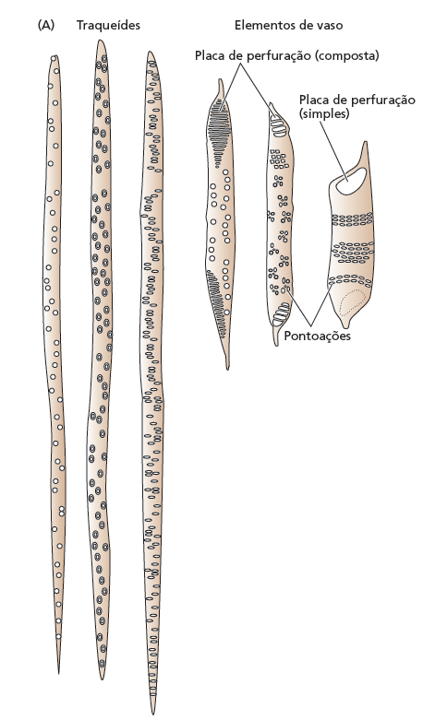
O xilema é constituído por células mortas altamente especializadas que, ao perderem seu conteúdo celular na maturidade, formam um sistema condutor oco e contínuo. O transporte de água e solutos ocorre através de dois tipos principais de células condutoras: os elementos de vaso e os traqueídes.
Os elementos de vaso, exclusivos das angiospermas, caracterizam-se por serem células cilíndricas, relativamente mais curtas e largas. Suas extremidades são conectadas por placas de perfuração, simples ou compostas, o que permite um fluxo longitudinal de água extremamente eficiente. Essa estrutura oferece baixa resistência hidráulica, assemelhando-se a canos conectados em série, garantindo o transporte de grandes volumes de água. A contrapartida dessa alta eficiência é a vulnerabilidade anatômica: sob elevada tensão hídrica, os elementos de vaso apresentam um risco substancialmente maior de sofrerem a formação de bolhas de ar, embolia ou cavitação.
Por outro lado, os traqueídes estão presentes em todas as plantas vasculares, desde as mais primitivas, como samambaias e gimnospermas, até as próprias angiospermas. Tratam-se de células muito mais alongadas e estreitas, com paredes celulares espessadas. Nelas, o fluxo hídrico ocorre de forma lateral, através de estruturas chamadas pontoações. Embora essa via imponha uma maior resistência ao fluxo, resultando em uma menor vazão por célula se comparada aos elementos de vaso, ela confere à planta uma altíssima segurança de transporte. O sistema de pontoações atua como uma barreira física que restringe a propagação de embolias, isolando o ar e protegendo a integridade da coluna d’água.
1.2.3 Movimento da água da folha para a atmosfera
O movimento final da água, transitando da folha para a atmosfera, é um processo dinâmico e estritamente regulado. Ao atingir o final do trajeto no xilema, a água líquida é descarregada nas células do mesofilo foliar.
A partir desse ponto, o percurso envolve uma mudança de fase conhecida como evaporação interna. A água passa da fase líquida para a fase gasosa (vapor) preenchendo os espaços intercelulares dentro da folha, com destaque para a câmara subestomática. O vapor de água, então, difunde-se por esses espaços buscando escapar para a atmosfera.
No entanto, o contato direto do vapor interno com o ar externo não é livre; o fluxo enfrenta duas barreiras físicas, denominadas resistências ao fluxo de vapor:
- Resistência Estomática Foliar: É a principal via de controle biológico da planta, sendo determinada diretamente pelo tamanho da fenda estomática (grau de abertura dos estômatos).
- Resistência da Camada Limítrofe: Refere-se à fina camada de ar estagnado que repousa imediatamente adjacente à superfície da folha, atuando como um obstáculo adicional à difusão do vapor para a atmosfera livre.
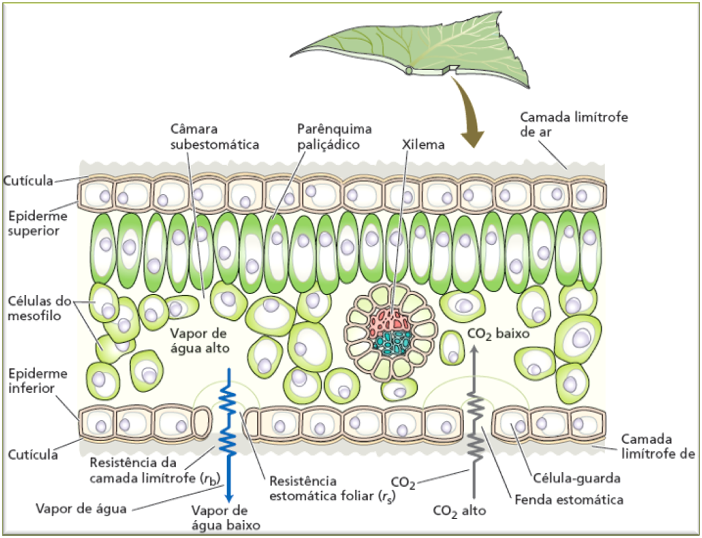
Os estômatos são estruturas microscópicas essenciais para a regulação das trocas gasosas entre a planta e a atmosfera. Eles ocorrem em altíssima densidade, frequentemente milhares por centímetro quadrado, e estão localizados predominantemente na epiderme inferior das folhas. Em algumas espécies, podem estar presentes na face superior em menor número e até mesmo em caules jovens, mas nunca ocorrem em raízes.
Cada estômato é composto por um poro central, delimitado e controlado por duas células-guarda altamente especializadas. O tamanho dessa abertura, que em sua amplitude máxima atinge fendas da ordem de 2 a 10 mícrons, é regulado unicamente pela pressão hídrica interna, o chamado controle por turgor.
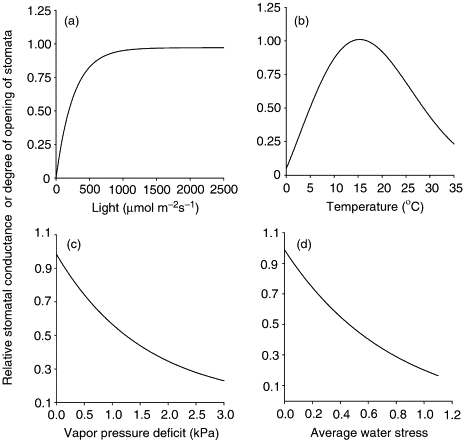
O controle da abertura e fechamento dos estômatos é governado por dois gatilhos principais que operam com propósitos distintos: a luz solar, que regula o ciclo diário normal de ganho de carbono, e o estado hídrico, que atua como um sistema de segurança contra a desidratação.

Em condições de umidade adequada no solo, a luz solar é o estímulo para a abertura estomática. Esse mecanismo dita o padrão diurno da condutância foliar, que acompanha estritamente a curva de incidência de radiação fotossinteticamente ativa ao longo do dia.
Fisiologicamente, o processo funciona via alteração ativa do potencial osmótico. A presença de luz estimula o bombeamento massivo de íons potássio (K+) para o interior das células-guarda. Esse forte acúmulo de solutos “puxa” a água das células vizinhas do mesofilo por osmose, elevando substancialmente a pressão de turgor interno. As células-guarda incham, curvam-se e abrem a fenda, permitindo a livre entrada de CO2 . Durante a noite, sem o estímulo luminoso, o potássio sai das células, a água acompanha o gradiente, o turgor cai e o estômato fecha, evitando perdas noturnas de vapor de água.
O cenário muda quando a umidade no solo se torna limitante. Se a demanda evaporativa da atmosfera for maior do que a capacidade das raízes de extrair e repor a água, a planta emite um sinal de alerta de estresse hídrico.
Nesta situação, o gatilho hídrico entra em ação e se sobrepõe imediatamente ao estímulo da luz. O sinal de alerta força a rápida saída dos íons de potássio das células-guarda, mesmo que a folha esteja sob sol pleno. A saída dos íons arrasta a água para fora, causando uma queda abrupta de turgor e induzindo o fechamento estomático diurno precoce.
Embora esse fechamento forçado durante o dia evite o colapso hidráulico e a dessecação severa dos tecidos vegetais, a consequência imediata é o bloqueio da entrada de carbono. É essa interrupção na fotossíntese promovida pelo fechamento estomático defensivo que explica as perdas de rendimento no campo durante as estiagens.
1.3 Evapotranspiração Relativa e Queda de Produtividade
O fechamento estomático atua como um mecanismo de sobrevivência imediato frente ao estresse hídrico. Contudo, essa estratégia de defesa tem um custo agronômico elevado: ao restringir a perda de água, a planta inevitavelmente bloqueia a entrada de CO2, paralisando a fotossíntese. Em termos práticos, o que salva a planta no curto prazo é exatamente o que penaliza a sua produtividade no final do ciclo.
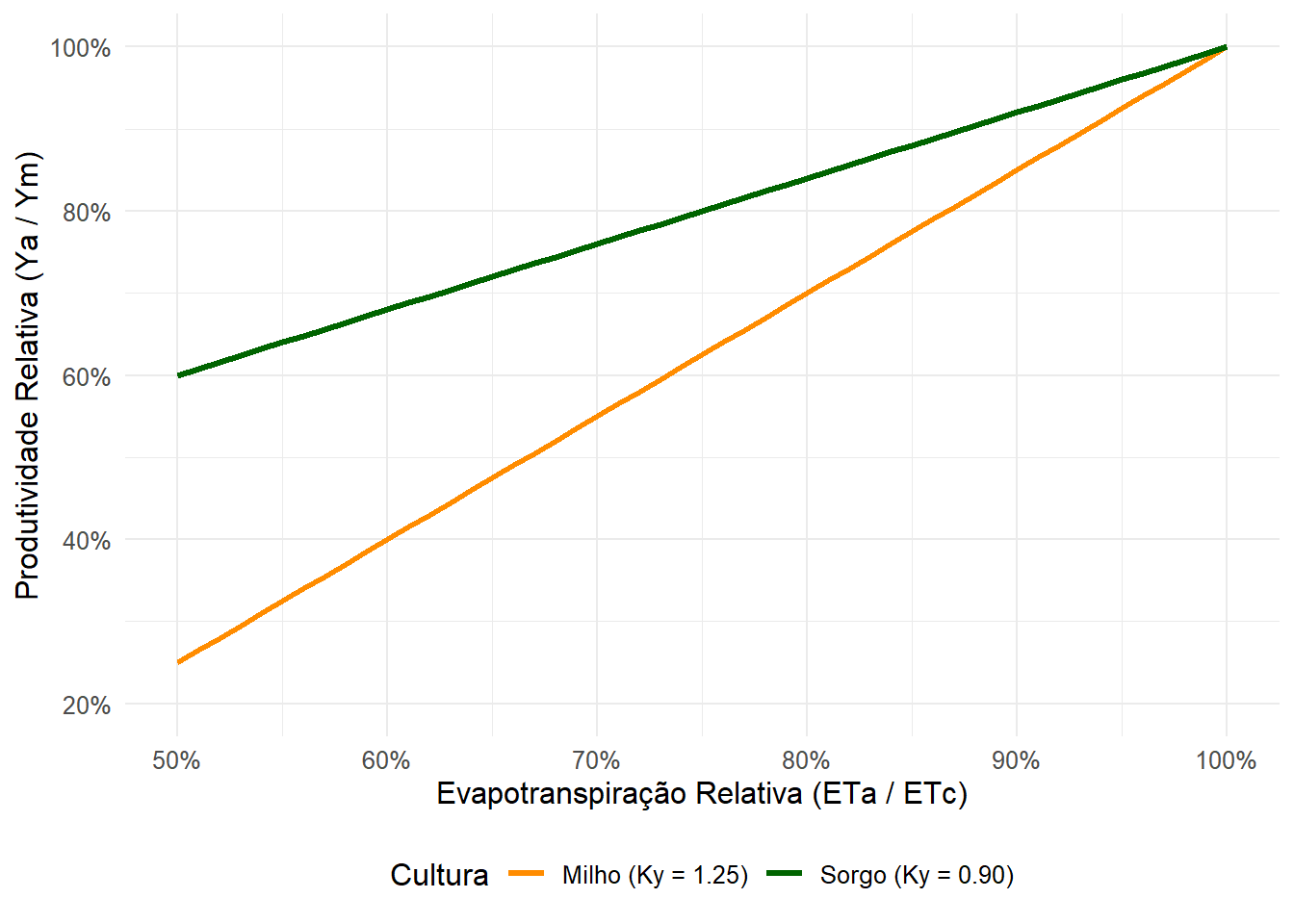
Para quantificar matematicamente a magnitude dessa penalidade, utilizamos a consagrada relação empírica proposta por Doorenbos & Kassam (1979)1, publicada no Boletim FAO 33. Essa equação relaciona a queda relativa de produtividade com o déficit hídrico relativo (ou evapotranspiração relativa) sofrido pela cultura:
\[ 1 - \frac{Y_a}{Y_m} = K_y \left( 1 - \frac{ET_a}{ET_c} \right) \tag{1.1}\]
Em que cada termo da equação representa os seguintes parâmetros ecofisiológicos e agronômicos:
- Ya: Produtividade real alcançada pela cultura no campo (kg/ha).
- Ym: Produtividade máxima potencial da cultura, obtida quando não há restrição hídrica ou nutricional (kg/ha).
- Ya/Ym: Rendimento relativo da cultura (uma fração ou porcentagem do potencial).
- ET_a: Evapotranspiração real da cultura sob as condições de umidade disponíveis no solo (mm).
- ETc (ou ETm): Evapotranspiração potencial ou máxima da cultura em condições padrão, sem estresse hídrico (mm).
- ETa/ETc: Evapotranspiração relativa.
- Ky: Fator de resposta da cultura ao déficit hídrico.
O grande trunfo desta equação é o fator Ky, que quantifica a sensibilidade específica de cada espécie (e fase fenológica) à falta de água. Ele atua como um multiplicador do impacto do estresse:
- Ky > 1: Indica uma cultura altamente sensível. A queda de produtividade é proporcionalmente maior do que a redução no consumo de água. (Exemplo: Milho, onde um pequeno déficit hídrico no pendoamento gera perdas severas de espigas).
- Ky < 1: Indica uma cultura mais tolerante. A planta consegue suportar certas restrições hídricas com perdas de produtividade proporcionalmente menores. (Exemplo: Sorgo ou Algodão).
- Ky = 1: A quebra de safra é diretamente proporcional ao déficit de água.
Compreender essa equação é essencial para o planejamento eficiente da irrigação, permitindo ao agrônomo prever quebras de safra e alocar a água disponível para os momentos em que a cultura é mais sensível (maior Ky).
Doorenbos, J. and Kassam, A.H. (1979) Yield response to water. FAO Irrigation and Drainage, Paper 33, Rome, 193 p.↩︎